

 |
|
دیانای
فهرست مندرجات
- واژهشناسی
- پیشینهی تاریخی کشف
- ساختار و ویژگیها
- کارکرد زیستی
- يادداشتها
- پيوستها
- پینوشتها
- جُستارهای وابسته
- سرچشمهها
- پيوند به بيرون
زیستشناسی مولکولی ژنتیک یاختهای
دِنــا یا دیانای (به انگلیسی: ، با تلفظ آمریکایی: )، سرواژهی عبارت دیوکسیریبونوکلئیکاسید (به انگلیسـی: )، نوعی اسـید نوکلئیک میباشد که دارای دستورالعملهای ژنتیکی است که برای کار کرد و توسعه بیولوژیکی موجودات زنده و ویروس مورد استفاده قرار میگیرد. نقش اصلی مولکول دیانای ذخیرهسازی طولانی مدت است.
| ▲ | واژهشناسی |
واژهی «دیانای» ()، مخفف (شیمی - زیستشناسی) «دیوکسیریبونوکلئیک اسید» یا «اسید دیاکسیریبونوکلیک» میباشد که به کوچکترین واحد وراثت اطلاق میشود و برابرنهادهی این واژه در زبان فارسی، «دِنا» است. بنابراین، دیانای، یکی از ماکرومولکولهای سلولی است که حامل اطلاعات وراثتی بوده و طی همانندسازی ژنتیکی از یک نسل به نسل بعد منتقل میشود و در داخل سلول از روی آن «آرانای» ( یا اسید ریبونوکلئیک) و پروتئین ساخته میشود.
| ▲ | پیشینهی تاریخی کشف |
کشف مادهای که بعدها دنا (دیانای) نام گرفت در سال ۱٨٦۹ بهوسیله «فردیک میشر»، پزشک سویسی، انجام شد. این دانشمند هنگام مطالعه بر روی گویچههای سفید خون، هستۀ سلولها را استخراج کرد و سپس بر روی آن محلول قلیایی ریخت. حاصل این آزمایش، رسوب لزجی بود که بررسیهای شیمیایی آن نشان داد، ترکیبی از کربن، هیدروژن، اکسیژن، نیتروژن و درصد بالایی از فسفر میباشد. میشر این ماده را «نوکلئین» نامید. زمانی که ماهیت اسیدی این ماده مشخص گردید، نام آن به «اسید دزاکسی ریبونوکلئیک» تغییر یافت.
با این وجود، عملکرد این مادهی شیمیایی که تنها در هستهی سلولها یافت میشود ناشناخته بود. پس از گذشت سالها، در سال ۱۹٣٠ کاسل و لوین دریافتند که دیانای حاوی فسفات، نوعی شکر بهنام «دیاکسی ریبوس» و چهار ترکیب متفاوت بهنام پایههای «نوکلئوتید» است که از سه قسمت تشکل شده است. یک قند پنتوز، یک گروه فسفات و از یکی از چهار باز آلی نیتروژندار حلقوی آدنین (A)، گوانین (G)، سیتوزین (C) و تیمین (T) تشکیل شده است.
شاید بزرگترین موفقیت پزشکی قرن بیستم، کشف ساختار دیاوکسی ریبونو کلوییک اسید (دیانای - مبنای مولکولی وراثت) باشد. دستیابی به دانش ساختار شیمیایی دیانای به دانشمندان اجازه داد که برای اولینبار دریابند چگونه دیانای تکثیر میشود و اطلاعات ژنی را از یک نسل به نسل بعدی انتقال میدهد. این کشف بسیار بزرگ، تقریباً بسیاری از جنبههای علم پزشکی را متحول کرد و پیشرفت چشمگیری در داروها، هورمونها و مواد مفید دیگر که بهلحاظ ژنتیکی مهندسی شده بودند، ایجاد نمود. این کشف بهتغییراتی بنیادین منجر شد.
در سال ۱۹۴۴ پزشک و باکتریشناس آلمانیتبار کانادایی مقیم آمریکا، اسوالد تیآوری و همکارانش، طی مجموعهای آزمایش روی باکتری، پی بردند که دیانای اطلاعات ژنتیک را انتقال میدهد. قبل از کار موفقیتآمیزآوری، بسیاری از بیوشیمیدانها عقیده داشتند که پروتئینها منبع اطلاعات ژنتیکی هستند. او، نشان داد که اسیدهای نوکلئیک دارای قند، اسید فسفریک و چند باز نیتروژندار میباشند. اندکی بعد مشخص شد که قند موجود در اسیدهای نوکلئیک میتواند ریبوز یا دئوکسیریبوز باشد. پس، اسیدهای نوکلئیک به دو دسته «دیانای» (DeoxyriboNucleic Acid) – که قند موجود در آنها دئوکسی ریبوز است – و «آرانای» (RiboNucleic Acid) - که قند موجود در آنها ریبوز است – تقسیم میشوند.
در سال ۱۹۴۸ لینوس پاولینگ کشف کرد که بسیاری از مولکولهای پروتئینی بهشکل یک مارپیچ هستند، و کموبیش شکلی همانند فنر دارند. در سال ۱۹۵۰، نیز اروین شارگاف نشان داد که اگرچه آرایش بازهای موجود در ساختار دیانای بسیار گوناگون است، ولی همواره نسبت باز آدنین و باز تیمین موجود در آن با هم برابر است و همینطور نسبت باز سیتوزین با باز گوانین. این دو یافته نقش مهمی را در آشکار شدن ساختار مولکول دیانای داشتند.
در دهه ۱۹۵٠، همچنان رقابت برای یافتن ساختار دیانای ادامه داشت و دو گروه از دانشمندان شدیداً بهدنبال کشف ساختار دیانای بودند. یکی از این گروهها در آزمایشگاه کاوندیش کمبریج در انگلستان مشغول بهکار بود و گروه دیگر در یک کالج سلطنتی در لندن. در دانشگاه کمبریج، فرانسیس کریک و جیمز واتسون برپایه کارهای پاولینگ کوشش داشتند تا با ارایه مدلهای فیزیکی ساختارهای احتمالی ممکن برای دیانای را محدود کنند تا سرانجام به ساختار درست دست یابند. گروه دیگر که در برگیرنده یک فیزیکدان بهنام موريس ویلکینز و یک فارغالتحصیل فیزیک بهنام ریموند گوسلینگ بود، همزمان سرگرم مطالعه دیانای بودند. این دو در سال ۱۹۵۱ با روزالیند فرانکلین که متخصص کریستالوگرافی اشعهی ایکس بود، در زمینه دیانای همکاری خود را آغاز کردند. آنها کوشش داشتند تا با روش آزمایشگاهی بهویژه با بهکارگیری تصاویر پراش اشعهی ایکس از مولکول دیانای، ساختار آن را معین کنند. بههر حال، روش کار این گروه با گروه پیشین متفاوت بود.

فرانسیس کریک (چپ) و جیمز واتسون با مدل اولیه مولکول دیانای تقریباً در همان زمانی که این کشف را به دنیا اعلام کردند
با این وجود، در سال ۱۹۵٠، ویلکینز که نمونهای خالص و بینظیر از دیانای را از فیزیکدانی سویسی دریافت کرده بود، با این نمونه، او توانست رشتههای دیانای را با یک میلهی شیشهای جدا کند. در همانسال، ویلکینز و گوسلینگ از طریق اشعهی ایکس، از این رشتهها عکس گرفتند و فرانکلین که در سال ۱۹۵۱ به آزمایشگاه آنها ملحق شده بود، نیز این کار را انجام داد.
اما یک سوءتفاهم باعث شد فرانکلین و ویلکینز، هر یک به تصور اینکه خودش مسئول کریستالوگرافی اشعهی ایکس بوده است، از ادامهی همکاری امتناع کنند. فرانکلین در سال ۱۹۵٢ از تیم خارج شد و تمامی عکسهای انکسار اشعهی ایکس را به ویلکینز تحویل داد. یکی از این تصاویر نشان داد که مولکول دیانای بهشکل مارپیچ دوبل است؛ ساختاری که به نردبانی پیچخورده شباهت دارد.
در همان زمان، زیستشناس آمریکایی، جیمز واتسن که در یک نشست در ناپل ایتالیا حضور یافته بود، یکی از عکسهای انکسار اشعهی ایکس ویلکینز را دید. او بلافاصله فکر کرد که مولکول میتواند یک مارپیچ دوبل (دوگانه) باشد. وی در پاییز ۱۹۵۱ به گروه دانشمندان آزمایشگاه کاوندیش پیوست و در آنجا توانست بیوفیزیکدان بریتانیایی، فرانسیس کریک را متقاعد کند که ترکیبی از مدلسازی - با استفاده از توپهای پلاستیکی، سیمها و صفحات فولادی - و روش کریستالوگرافی با اشعهی ایکس، میتواند آنها را به ساختار دیانای هدایت کند.
مارپیچ دوبل بهخودی خود تنها راز مولکول دیانای نبود. ساختار شیمیایی کلی آن هم به توضیح نیاز داشت. واتسن، بدون اطلاع از دستاوردهای ویلکینز، اینک متقاعد شده بود که دیانای ساختار مارپیچی دارد. او همراه با کریک، بهشدت در حال کار کردن روی تهیهی مدل پیچیدهی مولکول بود و توانست آن را در دومین هفتهی سال ۱۹۵٣ به پایان برساند. این مدل تمامی عناصر شیمیایی شناختهشده دیانای را با یکدیگر ترکیب کرده و تا حد زیادی بر الگوی انکسار مشاهده شده در عکس ویلکینز منطبق بود. واتسن و کریک بهدرستی نتیجهگیری کردند که دو رشتهی مارپیچ دوبل، که قبل از تقسیم سلولی از هم جدا میشوند، قالبها یا الگوهایی را برای تولید دو مولکول جدید همسان با مولکول اصلی ایجاد میکنند.
واتسن و کریک یک کپی از دستنوشتههای خود را برای ویلکینز فرستادند. این دستنوشتهها حاوی آن چیزی بود که ویلکینز و فرانکلین قبلاً انجام داده بودند و ویلکینز فکر میکرد محصول کار خودش است. ویلکینز پس از مطالعهی دستنوشتهها، نامهای برای آنها نوشت که اینگونه شروع میشد: «من فکر میکنم شما دو فریبکار هستید...» اما بهزودی بین آنها تفاهم ایجاد شد.

عکس معروف به تصویر ۵۱ کار روزالیند فرانکلین
در سال ۱۹۵۱، فرانکلین دریافت که دیانای با نگرش به میزان نم هوای پیرامون، میتواند دو شکل متفاوت داشته باشد و بنابراین نتیجهگیری کرد که بخش فسفات مولکول در سمت بیرونی آن قرار دارد. اندکی بعد او با بهکارگیری تصاویر اشعهی ایکس فهمید که دیانای در حالت «نمناک» از همگی ویژگیهای یک مارپیچ برخوردار است؛ این احتمال که حالت دیگر مولکول دیانای نیز به شکل مارپیچی باشد به ذهن او خطور کرد، ولی نمیخواست تا زمانی که شواهد پایانی برای این حدس پیدا کند، آن را اعلام نماید. در ژانویه ۱۹۵۳ ویلکینز که از به نتیجه رسیدن تحقیقات ناامید شده بود، نتایج تحقیقات فرانکلین را بدون اطلاع و خشنودی او، با واتسون در میان گذاشت. واتسون و کریک با بهکارگیری این نتایج مدلی بسیار شگفتانگیز را برای ساختار دیانای پیشنهاد نمودند. آنها مولکول را بهگونه دو زنجیر مارپیچی در برگیرنده نوکلئوتیدها تصور کردند که یکی از آنها بالا میرفت و دیگری پایین میآمد. کریک که بهتازگی یافتههای شارگاف را هم مطالعه کرده بود کوشش کرد با بهکارگیری آنها روش قرار گرفتن بازها را در مولکول دیانای مشخص کند. او اظهار کرد که بازها در میانه این مارپیچ دوتایی دو به دو به هم متصل میشوند تا فاصله میان دو مارپیچ ثابت بماند. آنها ادعا کردند که هر یک از این دو مارپیچ مولکول دیانای میتواند بهنام قالبی برای ایجاد دیگری بهرهگیری شود. در تقسیم سلولی این دو رشته از هم جدا میشوند و بر روی هر یک از آنها یک نمونه جدید همانند رشته مقابل پیشین ساخته میشود. با این روش بدون اینکه ساختار دیانای عوض شود، یک دیانای همانند آن فرآوری میشود. در اندک مواردی که در این روند خطایی پیش بیاید، گواه «جهش» خواهیم بود. مدل آنها چنان با اطلاعات برآمده از آزمایشها مطابقت داشت که بیدرنگ مورد قبول همه واقع شد. کشف ساختار دیانای را میتوان مهمترین یافته زیستی در صد سال گذشته دانست.
در ٢۵ آوریل ۱۹۵٣، مجلهی نیچر یک مقاله از آزمایشگاه کاوندیش کمبریج و دو مقاله از کالج سلطنتی لندن را با موضوع ساختار مولکولی دیانای چاپ کرد. ده سال بعد، در سال ۱۹٦٢، ویلکینز، واتسن و کریک بهطور مشترک، برندهی جایزهی نوبل فیزیولوژی و پزشکی شدند. بسیاری عقیده دارند کشف آنها تا حد بسیار زیادی مدیون کارهای «رزالین الیس فرانکلین» است. اما هنگامی که جایزه نوبل پزشکی به آنها تعلق گرفت، هیچ صحبت یا اشارهای در مورد کارها و تحقیقات رزالین فرانکلین به میان نیاوردند. در حالی که ۴ سال قبل، رزالین فرانکلین جوان پس از سالها تلاش در راه علم زیستشناسی به علت قرار گرفتن بهطور مستقیم در معرض اشعهی ایکس و کریستالوگرافی مبتلا به سرطان تخمدان شد و سرانجام در سال ۱۹۵۸ در ۱٦ آوریل در چلسی بعد از دو سال دستوپنجه نرمکردن با سرطان و در ۳۸ سالگی درگذشت. واتسون در کتاب مشهورش - مارپیچ دوگانه - نه تنها تقدیری از این دانشمند به عمل نیاورد، بلکه با عباراتی موهن از حضور فیزیکی وی یاد کرد که «او یک زن سردمزاج بدلباس بینمک بوده است»! او میگوید: «اگر من و کریک در ماه فوریه به نتیجه نرسیده بودیم، احتمالاً فرانکلین معما را حل میکرد؛ اگر رزالین به فکر کردن به دیانای ادامه میداد و این کار را رها نکرده بود، میتوانست جواب را پیدا کند.»

واتسون و کریک با مدل دیانای، بیست و یک سال بعد از کشف
کشف ساختار دیانای - همانند کشف اشعهی ایکس در اواخر قرن نوزدهم یا کشف باکتری، در بیش از دو قرن قبل - در علم پزشکی تغییرات بنیادی ایجاد کرد و افقهای ناشناختهای را فراروی این علم گشود. محققان امیدوارند با مجهزشدن به نقشهای از ژنومهای انسانی (کدهای کامل ژنتیکی)، در قرن آینده بتوانند دلایل ژنتیکی بسیاری از بیماریهای وراثتی از قبیل اسکیزوفرنیا، فیبروز کیستیک، هموفیکی و انواع سرطانها را ریشهکن کنند. مهمتر از همه اینکه بهاعتقاد بسیاری از دانشمندان، پیشرفت در ژنتیک مولکولی، زمینه را برای تغییرات اساسی در تشخیص و درمان بیماریها فراهم میآورد. بنابراین، بهجای درمان صرف علائم بیماریها، یعنی کاری که هماکنون انجام میشود، پزشکان هزارهی جدید ممکن است این توانایی را داشته باشند که قبل از بروز نشانهی بیماریها، دلایل آنها را تشخیص دهند و بر طرف کنند
با اینحال، با وجود موفقیتهای خیرهکنندهی پزشکی مغرب زمین در مبارزه با بیماریها که موجب افزایش طول عمر بشر شده است، وضعیت سلامتی کشورهای در حال توسعه رو به بدترشدن نهاده است. اگرچه واکسنها و پادزیستهای جدید میتوانند هر سال جان میلیونها انسان را در سرتاسر دنیا نجات دهند؛ اما همچنان افراد زیادی به مالاریا، سل، فلج اطفال، ذاتالریه و اختلالات دیگری که بهراحتی قابل درمان هستند، مبتلا میشوند. هماکنون افراد زیادی نیز با سندرم نقص ایمنی اکتسابی (ایدز) روی زمین زندگی میکنند، در حالیکه برای ادامهی حیات، به داروهای گرانقیمت دسترسی ندارند. یکی از بزرگترین چالشهای قرن آینده، یافتن راههایی است که دسترسی بهدستاوردهای عظیم پزشکی غرب را برای افرادی که بیشتر به آن نیاز دارند، امکانپذیر سازد.
دانشمندان امیدوارند در هزارهی جدید، بهنقشهی کاملی از کدهای ژنتیکی انسان دسترسی پیدا کنند که به آنها کمک خواهد کرد دلایل ژنتیکی همهی بیماریهای وراثتی را ردیابی کنند و داروهای ممکن را برای پیشگیری از آنها بیابند.
| ▲ | ساختار و ویژگیها |
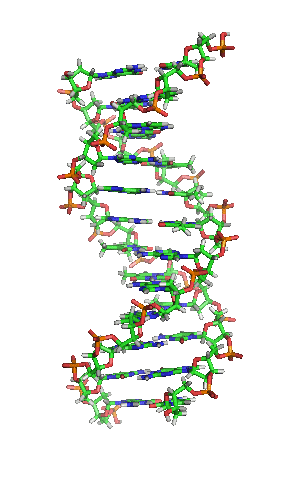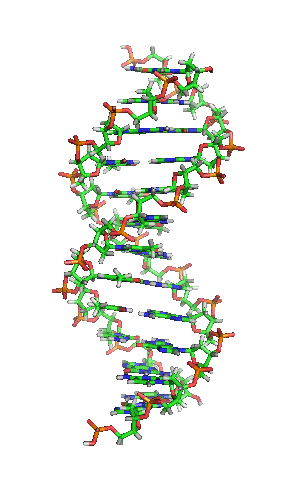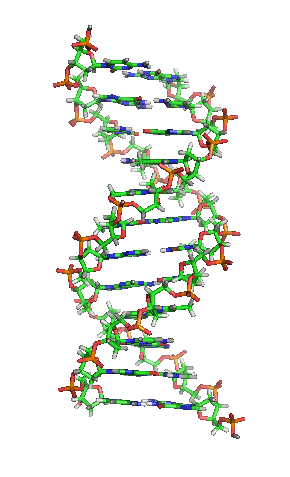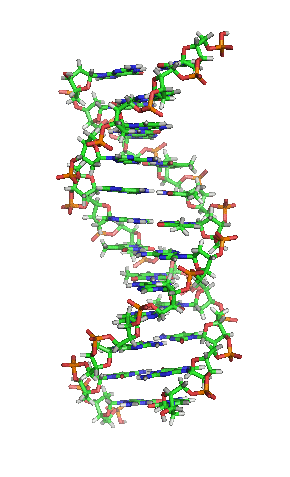 |
با روشن شدن حمل اطلاعات وراثتی توسط دنا، توجه محققان بهبررسی آن معطوف گرديد. تعيين ساختمان اوليه و سه بعدی اين مولكول تا حدی چگونگی حمل اطلاعات و همانندسازی كروموزوم را نشان داد. ابتدا اين نگرانی وجود داشت كه شايد مولكول دنا ساختمان بسيار پيچيده و غير عادی داشته باشد؛ اما بهزودی معلوم شد كه اين مولكول بهصورت مارپيچ مضاعف بوده و ساختمان سه بعدی ژنها مشابه يكديگر است و تنها تفاوت آنها در مورد تعداد و توالی نوكلئوتيدهای تشكيلدهنده يك ژن میباشد.
امروزه ملاحظه میشود كه ساختمان دنا چندان هم ساده نيست، بهطور مثال، دنا بعضی ويروسها تك رشته است (برخلاف اكثر مولكولهای ديگر كه بهصورت مارپيچ مضاعف یا مارپيچ دوگانه میباشند). ضمناً علیرغم اينكه مولكول دنا عمدتاً بهصورت مارپيچ راستگرد است، ولی بعضی از مناطق آن چپگرد میباشند. افزون بر این، همه مولكولهای دنا خطی نيستند و برخی حلقهای اند، از طرف ديگر در اثر تاب خوردن مولكولهای دنا بهدور پروتئينهای هستهای خاصی اشكال پيچيدهای بهنام ابرمارپيچ بهوجود میآيند كه ممكن است اين تاب خوردگیهای ثانويه، ساختمان مارپيچ مضاعف را بهگونهای تغيير دهند كه سبب باز شدن دو رشته فوق و بهوجود آمدن نواحی تك رشتهای گردد.
ابتدا تصور میشد كه ساختمانهای پيچيدهتر دنا اشكال فرعی هستند ولی بهزودی مشاهده گرديد كه فراوانی اين ساختمانها بسيار بيشتر از حدی است كه فرعی تلقی شوند. بنابراين ساختمان دنا نه تنها بهصورت نسبتاً ساده مارپيچ مضاعف نيست بلكه اشكال ساختمانی پيچيدهتری بر روی آن میتوانند بهوجود آيند و با درك علت ايجاد چنين ساختمانهايی میتوان تا حدی به نقش دنا در كنترل حيات پی برد.
بههر حال، دیانای (دئوکسی ریبو نوکلئیک اسید)، مولکولی است که دستورهای ژنتیکی مورد استفاده در توسعه و عملکرد تمام موجودات زندهٔ شناخته شده و بسیاری از ویروسها را کُدگذاری میکند. این مولکول اسید نوکلئیکی، شامل پروتئین و کربوهیدراتهاست که مانند همه اسیدهای نوکلئیک، از سه ماکرو مولکول اصلی تشکیل شده که برای زندگی همهٔ گونههای شناختهشده ضروری است.
بیشتر مولکول دیانای، یک ساختار دو رشتهایی ی زیستی میباشد که بهصورت حلقهی دور هم پیچخورده و بهشکل یک مارپیچ دوگانه () در آمده و هر رشتهی آن، از هزارها ساخته شده است. در واقع، ۴ نوع نوکلئوتید، مونومرهاى تشکیلدهندهی مولکولهاى دیانای هستند. این نوکلئوتیدها متشکلاند از یک قند مونوساکاریدی بهنام () و یک گروه فسفات و چهار باز آلی: (A)، (G)، (C) و (T)؛ که پایههای نیتروژنی نیز گفته میشوند.
بنابراین، ساختار شیمیایی دیانای بهصورت پیوند مشخصی از دو دنباله خطی این ۴ نوع نوکلئوتید میباشد. که این اتصالها فقط بهصورت (A-T)، و (T-A)، و (C-G)، و (G-C) وجود دارند (ترتیب قرار گرفتن نوکلئوتیدها در ساختار دیانای طورى است که همیشه A در مقابل T و C به G قرار مىگیرد).

ساختار دیانای و آرانای





نوکلئوتیدها بهوسیلهٔ بهصورت زنجیرهای بههم متصل میشوند، که از محل قند یک نوکلئوتید با فسفات نوکلئوتید دیگر پیوند ایجاد کرده و ساختاری رشته بلند شبیه ستون فقرات () قند و فسفاتی را ایجاد میکنند.
به سخن دیگر، رشتههای دیانای، از واحدهایی متشکل از قند و گروه فسفات تشکیل شده است که بهصورت متناوب و تکراری در طول رشته قرار گرفتهاند. قند مورد استفاده در دیانای، دئوکسی ریبوز است که نوعی پنتوز (قند پنج کربنی) میباشد. در این رشتهها، قندها توسط گروههای فسفری به یکدیگر پیوند داده شدهاند.
به بیان سادهتر، دیانای، شکل دو راهپله مارپیچی را دارد که بهدور هم پیچیده و بالا رفتهاند و هر یک از این راهپلهها یک نرده و یک پله دارد. اجزای سازندهای این نرده، که به آن (ستون فقرات) میگویند، گروههای فسفاتی و قند است. فسفات بار منفی دارد و قندی که در دیانای موجود است، از نوع دئوکسیریبوز میباشد. پلههای این دو راهپله از چهار باز آلی (بازهای نوکلئوتیدی): آدنین (A)، گوانین (G)، تیمین (T) و سیتوزین (C) ساخته شدهاند. بین پلهها که روبهروی هم قرار دارند، یک انفصال کوچکی وجود دارد. بنابراین، این دو راهپله (دو رشتهی دیانای) در محل انفصال پلهها که اندازه آنها باهم برابر نیست، بهوسیلهٔ پیوند کوالانسی بهصورت زنجیرهای بههم پیوند میخورند.
پورینها به همراه پیریمیدینها سازنده بازهای آلی هستند که در ساختار دیانای بهصورت نوکلئوتید شرکت دارند. نیمی از بازهای آلی موجود در اسیدهای نوکلئیک پورین هستند که شامل آدنین و گوانین میشود. این بازها در ساختار دیانای از طریق پیوندهای هیدروژنی به پیریمیدین منتاظر خود متصل میشوند. سه باز آلی سیتوزین، تیمین و اوراسیل که در ساختار دیانای و آرانای پیدا میشوند، از مشتقات پیریمیدینی هستند. از لحاظ ساختار شیمیایی، بازهای پورينی دو حلقهای و بازهای پيريميدينی يك حلقهای میباشند.

ساختار شیمیایی دیانای؛ پیوند هیدروژنی، به شکل نقطهچین نشان داده شده است. در توالی ناموازی دو رشتهٔ دیانای، بهصورت قراردادی، جفت شدن رشتهٔ بالایی از سر '۵ به سر '۳ و جفت شدن رشتهٔ پایینی از سر '۳ به سر '۵ میباشد
بنابراین، در دیانای، نوکلئوتید هر رشته از طریق چهار باز آلی موجود در هر دو رشته به یکدیگر متصل میشوند. این اتصال بین دو باز آلی نوکلئوتیدهای دو طرف رشته میباشد به این بازهای متصل بههم باز مکمل گفته میشود. در این میان، باز آدنین مکمل تیمین، و باز گوانین مکمل سیتوزین میباشد. با این حال، دو نوکلئوتید با ترکیبات مکمل مخالف روی رشتههای دیانای، که با پیوند هیدروژنی بههم متصل شدهاند، «» () نامیده میشود.

توالی دیانای - در تصویر - یک الگوی جفت شدن دو رشتهای را نشان میدهد.
پیوند بازهای مکمل با یکدیگر از طریق پیوند بین هیدروژن یک باز با مولکول نیتروژن یا اکسیژن باز مکمل حاصل میشود. این پیوند از نوع قوی کووالانسی نمیباشد و در نتیجه بهراحتی شکسته میشود و قابل جایگزینی است. بههمین علت زنچیره دورشتهایی دیانای، بهراحتی در اثر فشار یا گرمای بالا از یکدیگر جدا میشوند. با این حال، ییوند مولکول هیدروژن بین دو باز مکمل آدنین-تیمین با گوآنین-سیتوزین متفاوت است. در گوآنین-سیتوزین سه مولکول هیدروژن پیوندی وجود دارد، در حالیکه در آدنین-تیمین دو مولکول هیدروژن پیوندی موجود است؛ در نتیجه میزان تعداد بازهای مکمل گوانین-سیتوزین تعیینکنندهی استحکام دیانای میباشد، بهطوریکه هرچه مقدار آن بیشتر باشد دیانای مستحکمتر است.
| ▲ | کارکرد زیستی |
دیانای یکی از ماکرومولکولهای زیستی میباشد که در انتقال دادههای ژنتیکی نقش دارد. این ماکرومولکولها، پلیمری از زیرواحدهای نوکلئوتیدی هستند.[*] کارکرد اصلی دیانای الگو بودن برای ساخته شدن یک اسید نوکلئیک دیگر بهنام آرانای است.
دیانای، متکاملترین مولکول ایجاد شده توسط طبیعت تلقی میشود که مشخصهی مهم این مولکول، هوشمندی آن است. دو رشته دیانای که از نظر توالی بازها، مکمل یکدیگر میباشند، میتوانند یکدیگر را شناسایی کرده، پیوند هیدروژنی برقرار کنند.
از نظر ژنتیک مولکولی، سه فرآیند اصلی در استفاده از اطلاعات ژنتیکی مطرح است: نخستین فرآیند، همانندسازی دیانای یا نسخهبرداری از دیانای مادر و تولید مولکولهای دیانای با توالیهای نوکلئوتیدی یکسان میباشد؛ دومین فرآیند سنتز آرانای از روی دیانای است، که طی آن، قسمتهایی از پیام ژنتیکی کُد شده در دیانای دقیقاً بهصورت آرانای، نسخهبرداری میشود؛ و سومین فرآیند، ترجمه میباشد که به موجب آن، پیام ژنتیکی کُد شده در آرانای پیامرسان () بر روی ریبوزومها به پلیپپتیدی با توالی مشخص از اسیدهای آمینه ترجمه میشود. در نتیجه، محصول نهایی دو فرآیند اخیر، معمولاً یک پروتئین است.
| ۱ | همانندسازی دیانای |
دیانای، در داخل یاخته، بخشی از سازهیی کروموزوم میباشد. در زمان تقسیم یاخته، از هر کروموزوم، دو نسخه ساخته میشود؛ که این فرایند تکثیر به دو نسخه را نسخهبرداری یا همانندسازی دیانای (DNA replication) مینامند.[٢٣] کروموزومها در جاندران «یوکاریوتی» مانند جانوران، گیاهان، قارچها، و آغازیان، در بخشی هسته یاخته قرار میگیرد، در حالی که در جاندران سادهتر «پروکاریوتی» مانند باکتری و آرکیها - که سلول آنها فاقد هسته است - در سیتوپلاسم یاخته قرار گرفته و جایگاه مشخصی ندارد. درون کروموزومها، پروتئینهای کروماتینی مانند «هیستونها» باعث متراکمشدن و سازماندهی دیانای میشوند. این ساختار متراکم، تعامل بین دیانای و سایر پروتئینها را هدایت میکند و به کنترل اینکه کدام بخشهای دیانای نسخهبرداری شوند، کمک میکند.
در همانندسازی دیانای، دو مولکول دیانای تولید میشود که هر یک، دارای یک رشتهٔ جدید و یک رشتهٔ قدیمی هستند (ردیف نوکلئوتیدها در هر یک از مولکولهای دیانای حاصل، یکسان است) که باعث میشود دیانایهای دختر دقیقاً مشابه دیانای مادر باشند. به این روش تکثیر که هر دیانای دختر از یک رشته از دیانای مادر و یک رشتهٔ نو ساخته شده است طریقهٔ نیمهحفظ شده میگویند.
در واقع، در همانندسازی دیانای، دو رشتهٔ پلینوکلئوتیدی دیانای به کمک آنزیم هلیکاز مانند زیپ از یکدیگر جدا میشوند و سپس از روی هر رشته، رشتهٔ جدیدی ساخته میشود؛ به این ترتیب که آنزیمی بهنام دیانای پلیمراز (DNA Polymerase) بر روی نوار پلی نوکلئوتیدی حرکت میکند و با استفاده از نوکلئوتیدهای آزاد که در سیتوپلاسم وجود دارند (دارای ۲ یا ۳ گروه فسفات هستند) هر نوکلئوتید را در مقابل نوکلئوتید مکمل خود قرار میدهد. باید در نظر داشت که در دو طرف هر رشته اعدادی گذاشته شده است که یک طرف '۵ و طرف دیگر '۳ است و در رشته مقابل هم برعکس رشته دیگری میباشد؛ یعنی یک طرف از یک رشته از هر دو طرف خود (عمودی-افقی) به عدد دیگر میرسد. مسیر همانندسازی هم همواره از '۵ به '۳ میباشد.
آنزیم دیانای پلیمراز توانایی دیگری نیز دارد و آن «ویرایش» است. در صورتی که نوکلئوتید اشتباهی به دیانایهای دختر اضافهشود، یعنی مکمل نباشد، این آنزیم بر میگردد و نوکلئوتید غلط را جدا و آن را با نوکلئوتید صحیح تعویض میکند. اگر این اشتباهات تصحیح نشوند در دیانایهای دختر باقی میماند و به نسل بعد منتقل میشود. به این اشتباهات تصحیح نشده «جهش» میگویند.
در این پروسه، ژن موجود در مولکول دیانای که قطعههایی از دیانای هستند، پیامهای ژنتیکی را باخود حمل میکنند. افزون بر این، این ژنها برای مواردی چون ساخت مولکولهای آرانای و پروتئین در یاخته (سلول)، نیز مورد استفاده قرار میگیرد.[٢٢]
| ٢ | رونویسی و برگردان دیانای |
دیانای، طی دو فرآیند «رونویسی» (Transcription) و «ترجمه» یا «برگردان» (Translation)، به پروتئین تبدیل میشود. در این روند، ژنها، بهعنوان کوچکترین واحد وراثت، نقش مهم دارند.
ژن واحد مولکولی وراثتِ یک ارگانیسم زنده است. بیشتر ژنها اطلاعات مربوط به ساخت پروتئینها را در بر دارند و معمولاً در توالیهای مولکول دیانای ذخیره میشوند. این توالی دورشتهای دیانای، غیر قطبی و نامحلول در آب است.
در واقع، در رشتههای دیانای، توالی چهار باز آلی باعث رمزگذاری رشته زنتیکی میشود که این رمزهای ژنتیکی ()، توسط مولکول آرانای در مرحله برگردان خوانده میشود و برای ساخت اسید آمینه که واحدهای سازنده پروتئین هستند، مورد بهرهجویی قرار میگیرد.
اسید آمینه واحد تشکیلدهنده پروتئین است. در طبیعت، بیش از بیست نوع اسید آمینه (۲۲ اسید آمینه) برای ساخت پروتئینها شرکت دارند که میتوانند با هر ترکیب و به هر تعداد در ساختار یک پروتئین دخالت کنند. اسیدهای آمینه در هنگام ساختهشدن پروتئینها از آرانای پیامرسان (یا ) به یکدیگر میپیوندند.
۱ رونویسی عمل رونوشتبرداری از دادههای نوکلئوتیدی دیانای و تبدیل آن به دادههای نوکلئوتیدی آرانای میباشد. به عبارت دقیقتر، رونویسی فرآیندی است که ضمن آن با دخالت آنزیم اختصاصی آرانای پلیمراز () و مصرف مولکولهای پرانرژی «آدنوزین تریفسفات» ( با کوتهنوشت ATP) و بهکارگیری نوکلئوتیدها، ترکیبهای نوکلئوتیدی موجود در مولکول دیانای به ساخت آرانایهای گوناگون شامل آرانای پیامرسان ()، آرانای حامل ()، آرانای ریبوزومی ()، و ... میانجامد.
در پروکاریوتها یک نوع آنزیم آرانای پلیمراز رونویسی همهٔ گونههای آرانای را انجام میدهد؛ اما در یوکاریوتها علاوه بر آنزیم آرانای پلیمراز ویژهای که در دو اندامک کلروپلاست و میتوکندری آنها موجود است، سه گونهٔ دیگری از آنزیم آرانای پلیمراز نیز در هستهٔ آنها موجود میباشد که آنها را با شمارههای ۱ و ۲ و ۳ مشخص میکنند:
- آرانای پلیمراز ۱ (RNA polymerase I) فقط رونویسی ژنهای آرانای ریبوزومی () را انجام میدهد.
- آرانای پلیمراز ۲ (RNA polymerase II) رونویسی پیشسازهای آرانای پیامرسان () و نیز برخی از آرانایهای بیرمز (Non-coding RNA) را انجام میدهد.
- آرانای پلیمراز ۳ (RNA polymerase III) به رونویسی ژنهای آرانای حامل () و نیز برخی از آرانایهای بیرمز کمک میکند.
بهطور کلی، دستگاه رونویسی یوکاریوتی پیچیدهتر از دستگاه رونویسی پروکاریوتی است. هر یک از آنزیمهای آرانای پلیمراز ۱، ۲ و ۳ کمی بزرگتر از آنزیم پروکاریوتی و از بیش از ده زیرسازه تشکیل شدهاند. همانندی چیدمان نوکلئوتیدها در ژنهای رمزگردان برخی از زیرسازهها نشان میدهد که این زیرسازهها با یکدیگر و حتی با زیرسازههای آنزیم پروکاریوتی ریشه تکاملی مشترکی دارند.
بههر حال، ساختهشدن آرانای از روی دیانای (رونویسی)، نخستین گام برای ساخت پروتئینها میباشد که در سه مرحله انجام میشود:
- مرحله آغاز (Initiation): رونویسی با پیوند آرانای پلیمراز به بخشی از ژن بهنام راهانداز ژن (Promoter) آغاز میشود. راهانداز، بخشی از دیانای است که به آرانای پلیمراز امکان میدهد رونویسی را از جای درستی آغاز کند و مثلاً این کار را از میان ژن آغاز نکند. راهانداز در نزدیکی جایگاه آغاز رونویسی (Transcription Initiation Site) جای دارد. جایگاه آغاز رونویسی، به نخستین نوکلئوتیدی از دیانآ گفته میشود که رونویسی میشود.
- مرحله ادامه (Elongation): آرانای پلیمراز، دو رشتهٔ دیانای را از یکدیگر باز میکند.
- مرحله پایانی (Termination): آرانای پلیمراز هم چون قطاری که روی ریل حرکت میکند، در طول نوکلئوتیدهای دیانای به حرکت در میآید و در مقابل هر یک از دئوکسی ریبوتوکلئوتیدهای دیانای، ریبونوکلئیوتید مکمل را قرار میدهد و همچنین، هر ریبوتوکلئوتید جدید را به ریبونوکلئوتید پیشین پیوند میدهد. در رونویسی نیز از همان قوانین جفت شدن بازها که در همانندسازی دیانای بهکار میرود، استفاده میشود. تنها تفاوت این است که در مقابل دئوکسی نوکلئوتید آدنیندار در دیانای، ریبونوکلئوتید یوراسیلدار، در آرانای جای میگیرد.
مرحله آغاز رونویسی (RNAP = آرانای پلیمراز)
مرحله ادامه رونویسی
مرحله پایان رونویسی
سپس، آرانای پلیمراز، دیانای و آرانای پیامرسان تازهای ساخته میشود و بعد از رونویسی جایگاه پایان رونویسی (Transcription Termination Site)، از یکدیگر جدا میشوند و مولکول آرانای پیامرسان برای مرحلهٔ بعدی یعنی رمزخوانی، آزاد میشود.
٢ در ترجمه (رمزخوانی) یا سنتز پروتئین، چیدمان نوکلئوتیدها در آرانای پیامرسان به چیدمان اسیدهای آمینه در پروتئین برگردانده میشوند. در این فرآیند، در واقع اسیدهای نوکلئیک که رمزهای یک ژن هستند در ریبوزوم (Ribosome) خوانده میشوند و از روی آن رمزها، پروتئین که از اسیدهای آمینه درست شده است، ساخته میشوند.
پروتئینها کارگران واقعاً سختکوش سلولها هستند. مثل دیانای و آرانای، آنها زنجیرههایی از مولکولهای کوچکتر متصل به یکدیگرند (یعنی مولکولهای تشکیلدهندهی پروتئین به آمینو اسیدها موسوماند). برخی پروتئینها مثل کراتین (Keratin) و کولاژن (Collagen) بافتها و قسمتهای مختلف بدن را در شکل خود نگاه میدارند. سایر پروتئینها، مثل انسولین، بهعنوان پیامرسان در بین سلولها یا مثل هموگلوبین بهعنوان تأمینکنندهی مواد لازم (در این مورد اکسیژن) و قراردادن آنها در جاهایی عمل میکنند که اگر وجود نداشتند، مواد مزبور به اندازهی کافی به سلولها نمیرسید. سایر پروتئینها، مثل ترانسکریپتیز (Transcriptase)، آنزهایمهایی هستند که تقریباً تمامی واکنشهای بدن را تنظیم میکنند.
برای اینکه پیامهایی زنتیکی آرانای پیامرسان (mRNA) خوانده و به پروتئین تبدیل شوند، آرانای پیامرسان باید به بیرون از هستهی یاخته مهاجرت کند و در محیط محلولی که هستهی یاخته را احاطه کرده است و سیتوپلاسم (Cytoplasm) نامیده میشود، قرار گیرد. در سیتوپلاسم، آرانایهای پیامرسان در اطراف سلول میچرخند تا سرانجام بهوسیلهی تشکیلات مولکولی بهنام ریبوزومها بهدام میافتند و پیامشان خوانده میشود و به پروتئین تبدیل میگردند.
بنابراین، فرآیند ترجمه در درونیاخته و توسط ریبوزومها صورت میگیرد. در رمزخوانی آرانایهای پیامرسان، آرانایهای حامل، آرانایهای ریبوزومی، آرانایهای بیرمز و... شرکت دارند. با این حال، وقتی یک مولکول آرانای پیامرسان به یک ریبوزوم متصل میشود، حروف ژنتیکی آن بهصورت هر سه حرف (سه نوکلئوتید) در یکبار خوانده میشوند. هر سه حرف، مثل یک کلمهی ژنتیکی است که معرف یک آمینواسید است (بیش از ٢٠ امینواسید برای انتخاب، و ٦۴ مجموعهی سه حرفی یافت میشوند). با خواندهشدن هر مجموعهی سه حرفی، ریبوزوم، آنقدر آمینواسید مناسب به زنجیرهی پروتئینی در حال رشد میافزاید تا با یک مجموعهی سه حرفی که دستور توقف میدهد، برخورد کند. به این ترتیب، تغییر آرایش پایهها در دیانای اصلی میتواند زنجیرهی آمینواسید پروتئین را تغییر دهد و موجب تغییر عملکرد آن شود. اینگونه تغییرات بهنام «جهش» یا «» () موسوم است. برخی جهشها مخربند؛ برای مثال انسولین معیوب موجب بروز دیابت میشود. هموگلوبین معیوب موجب بروز کمخونی میشود. برخی از آنها، تفاوتهای کوچکی را موجب میشوند و تعداد بسیار معدودی از آنها، موجب بهبود و بهدنبال آن موجب فراهم آمدن مادهی خام تکامل میشوند.
فرایند رمزخوانی
بههر حال، فرآیند ترجمه را نیز میتوان به سه مرحله آغاز، ادامه و پایانی تقسیم کرد:[*][*]
- مرحله آغاز: بخش کوچکتر ریبوزوم در مجاورت کدون آغاز (Start codon) به آرانای پیامرسان (mRNA) متصل میشود. سپس بخش بزرگتر ریبوزوم به بخش کوچک میپیوندد و ساختار ریبوزوم برای ترجمه کامل میشود. هر ریبوزوم دو جایگاه دارد: جایگاه اول، جایگاه P برای پلیپپتید در حال ساخت و جایگاه دوم، جایگاه A برای امینو اسید. در این مرحله، آرانای حامل (tRNA)، که ناقل متیونین است به جایگاه P وارد میشود و در آنجا با کدون آغاز (Start codon) رابطهی مکملی برقرار میکند.
- مرحله ادامه: رمز و پادرمز، ابتدا وارد جایگاه اول (جایگاه P) و رمز و پادرمز بعدی وارد جایگاه دوم (جایگاه A) میشوند. بعد از قرار گرفتن رمز دوم آرانای پیامرسان (mRNA) در جایگاه دوم، آرانای حامل (tRNA) مکمل به آن متصل شده و اسید آمینه جایگاه اول به اسید آمینهٔ جایگاه دوم پیوند پپتیدی میدهد و رمز آغاز از جایگاه اول خارج میشود. در این حال، ریبوزوم به اندازهٔ یک رمز بر روی آرآنای پیامرسان به جلو حرکت میکند و رمز و پادرمز دوم به جایگاه اول که به تازگی خالی شده است وارد میشوند و رمز سوم به جایگاه دوم وارد میشود. ریبوزوم به همین ترتیب بر روی آرانای پیامرسان حرکت میکند و تا انتهای آن ادامه میدهد تا به رمز پایان برسد.
- مرحله پایانی: وقتی یکی از رمزهای پایان درون جایگاه A قرار گیرد، ترجمه پایان میپذیرد. چون هیچ آرانای حاملی برای رمز پایان وجود ندارد و در این حال دو بخش ریبوزوم یعنی آرانای پیامرسان و پروتئین ساختهشده از یکدیگر جدا میشوند.
فرایند رونویسی و ترجمه
[▲] يادداشتها
[▲] پيوستها
علی رنجبران، مولکولهای دیانای از حیات روی زمین قدیمیترند؟
مهبد ابراهیمی، راز حیات
[▲] پینوشتها
واژهٔ مصوب فرهنگستان زبان و ادب فارسی، دفتر نخست تا چهارم، ۱۳۷۶ تا ۱۳۸۵
دیانای، از ویکیپدیا، دانشنامهٔ آزاد
اسوالد آوری (انگلیسی: Oswald Avery؛ زاده ۲۱ اکتبر ۱۸۷۷ - درگذشته ۲۰ فوریه ۱۹۵۵) یک دانشمند در زمینه زیستشناسی مولکولی و بیوشیمیدان آلمانیتبار اهل کانادا بود.
1 2 3 4 5 6 7 8 9
مولکول دیانای یک ساختار دو رشتهای متشکل از چهار پایه است. این پایهها عبارتند از آدنین، گوانین، سیتوزین و تیمین. نحوه کنار هم قرار گرفتن این چهار باز در دیانای ،آرایش ژنها را تعیین میکند. محققان، در سال ٢٠۱۱ میلادی، پایه دیگری در ساختار دیانای کشف کردهاند که ۵- فورمیل سیتوزین یا 5fC خوانده میشود. محققان هنوز نمیدانند که این پایه چه کارکردی در دیانای دارد؛ اما مشخص شدهاست که این پایه همواره در همه بافتهای زنده بدن موشها حضور دارد.
5fC پیش از این، تصور میشد که یکی از عواملی باشد که نحوه بیان ژنها در مولکول دیانای را تحت تاثیر قرار میدهد. تا کنون محققان فکر میکردند که 5fC تنها یک ماده موقتی در دیانای است و بعدا با عملکرد آنزیمها حذف میشود، اما هماکنون مشخص شده است که 5fC یک عامل موقتی نیست و حضور دائمی آن در بافتهای زنده نشان میدهد که نقش مهمی در مولکول دیانای دارد.گروهی از محققان دانشگاه کمبریج از روشهای اسپکتروفوتومتری با قدرت تفکیک بسیار بالا استفاده کردند تا سطوح 5fC را در بافتهای زنده موشهای بالغ و جنین موشها و همچنین سلولهای رویانی مورد بررسی قرار دهند. آنها دریافتند که 5fC در تمام این بافتها وجود دارد، هرچند که تشخیص آن با دشواری زیادی همراه است.این پایه در هر موقعیت ویژهای در ژنوم یافت میشود. مشخص شده است که در تمام بافتهای بدن میتوان این پایه را پیدا کرد هر چند سطوح آن بسیار اندک است.محققان میزان جذب کربن پایدار و ایزوتوپ هیدروژن را در 5fC سلولهای موش اندازه گرفتند. در صورتی که 5fC یک مولکول موقتی و ناپایدار بود، جذب ایزوتوپ به میزان زیادی صورت میگرفت، اما در عوض، آنها مشاهده کردند که هیچ جذبی در بافت مغز صورت نمیگیرد که نشان میداد 5fC یک عامل پایدار در ساختار دیانای است.این یافتهها در مجله Nature Chemical Biology منتشر شده است.
[▲] جُستارهای وابسته
□
□
□
[▲] سرچشمهها
□
□
□
□
[▲] پيوند به بیرون
□ [۱ ٢ ٣ ۴ ۵ ٦ ٧ ٨ ٩ ۱٠ ۱۱ ۱٢ ۱٣ ۱۴ ۱۵ ۱٦ ۱٧ ۱٨ ۱۹ ٢٠ ٢۱ ٢٢ ٢٣ ٢۴ ٢۵]
ردهها: │ زیستشناسی │ زیستشناسی مولکولی │ ژنتیک یاختهای








